会议回顾
2023年8月24日,广州实验室苗智超研究员邀请了Pascale Romby教授进行标题为“以金黄色葡萄球菌为例阐述致病菌中的调控RNA:新发现及未来的挑战”的线上学术报告。Pascale Romby教授是法国科学院UPR 9002 CNRS-ARN中心主任与Stras-RNA的负责人,其研究方向为大肠杆菌和金黄色葡萄球菌中结构mRNA的翻译起始及其在细菌中的调控机制的研究,条件病原体金黄色葡萄球菌中调控RNA及其网络的功能的研究等。
关于金黄色葡萄球菌的研究在很久以前就已经开展。总所周知,在细胞中,转录对感应与传输外界信号而言尤为重要。一般来说,一种蛋白质调节几个目标基因,而一个被调节的基因是几个调节因子的目标。在大肠杆菌中,大约有200至300种转录调节蛋白。然而,F.Jacob和J.Monod在1961年提出,“在信息传递过程中,结构信息必须由一个非常短暂的中间体携带,这个中间体迅速形成,也迅速被破坏...这是对环境变化的快速遗传反应所必需的特性。”如今我们了解到,由信使RNA执行的后转录和调控对于细胞的快速响应非常重要,尤其是细胞对不同环境进行的的新陈代谢过程。
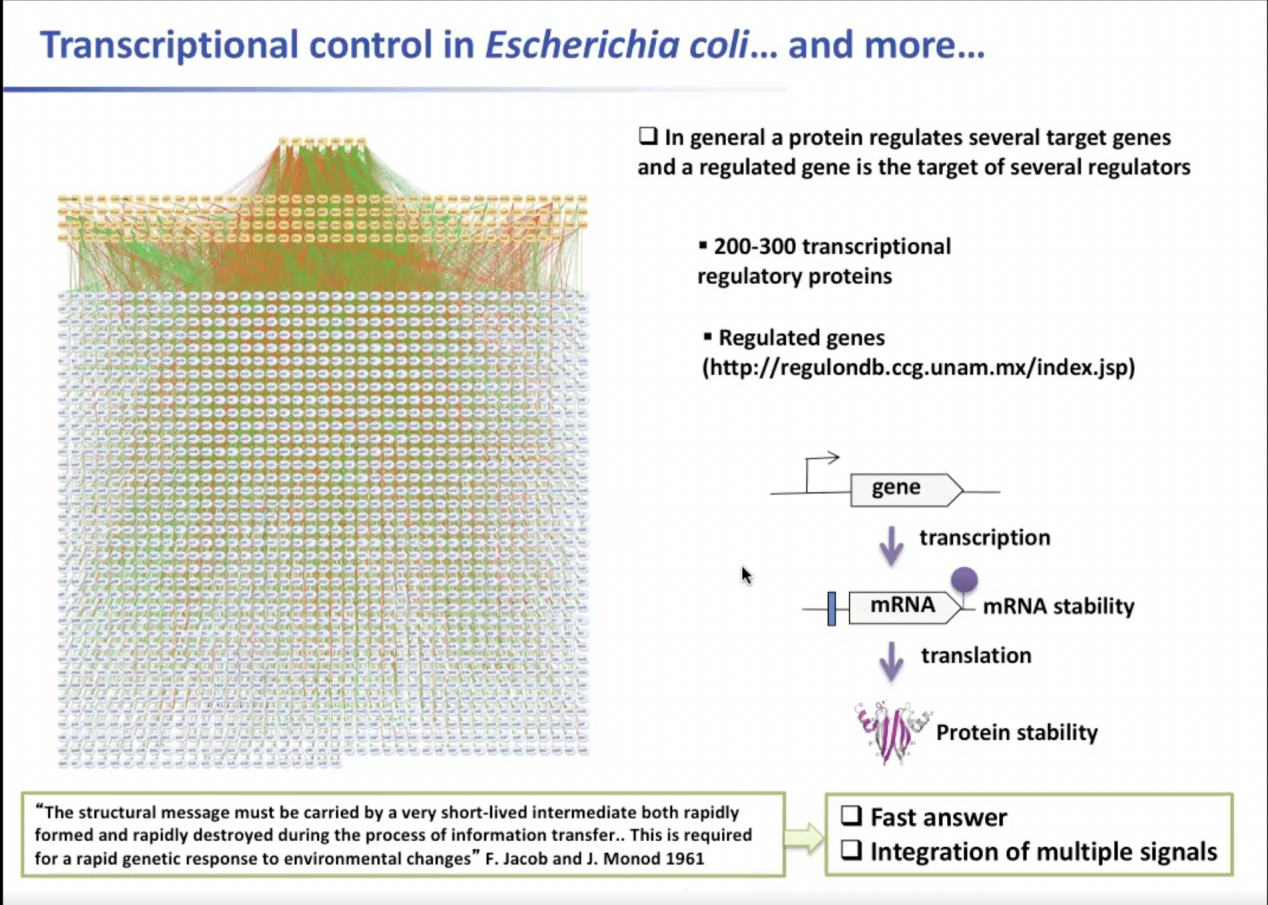
图1. 信使RNA执行的后转录和调控
以下是RNA分子在大肠杆菌中基因调控主要功能的例子。因为mRNA结构上有特定的基序,传感信使RNA(sensor mRNA)中一种名为热传感RNA(Thermosensor)的RNA分子序列中有一段碱基非典型配对(noncanonical pairing)的基序,其核糖体结合位点位于一段结构为“发夹弯(hairpin loop)”的结构中。这个结构会对温度升高迅速作出反应,“发夹弯”结构会变形,此时其结构可以在翻译过程中识别核糖体,因此这种RNA被称为热传感RNA。此外还有很多拥有反式作用配位体位点的代谢物分子感应核糖开关,一些配位体也可以是核糖开关。很多mRNA的5’非翻译区(5’UTR)由特定的酶解链并在核苷酸,维他命与氨基酸等合成物发生作用。作为蛋白调控元件,我们知道很多RNA与蛋白质结合,可据此识别特定RNA结构调控RNA行为。另外,在许多细菌的基因组中小非编码RNA(sncRNA)都有表达,它的主要作用是作为反义RNA与不同区域的mRNA结合,抑制许多mRNA的合成,或对蛋白质进行隔离。但是一些小非编码RNA也会隔离蛋白质。mRNA可被视作小非编码RNA的储存库,因为许多mRNA携带更长的5 ' 3 ' 非翻译区,不能被加工从而产生小非编码RNA,这些小非编码RNA会作用于其他信使RNA。因此实际情况高度复杂。因为有很多反义RNA功能与mRNA重叠,且mRNA可以与其他mRNA相互作用,许多mRNA也受到反义RNA的调控。总而言之,一个RNA分子在基因调控功能上可能有多种功能。
因此,一个关键的问题是如何解码mRNA的调控信号。在mRNA上找到这样的标记并猜出调节mRNA的反义配体是什么并不容易。此外,我们知道许多小非编码RNA,还有热传感RNA和一些核开关也可以与蛋白质一起作用。
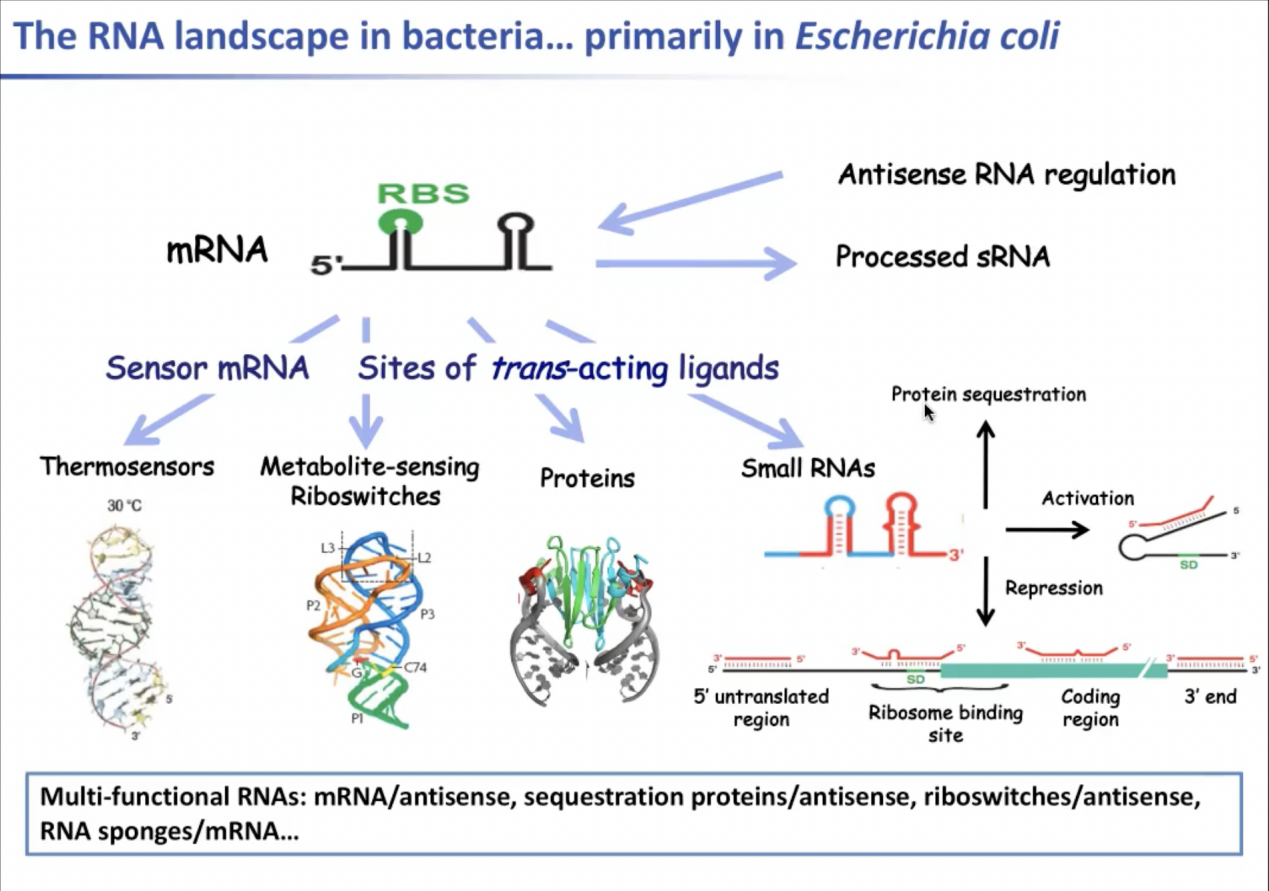
图2. RNA分子在大肠杆菌中执行调控功能
比如在大肠杆菌中,我们知道Rho蛋白是一种参与亚mRNA转录的终止及许多mRNA的调控的蛋白质。这些mRNA是由一些小的非编码RNA调控的。而这些蛋白质是大肠杆菌所必需的。许多核糖核酸酶也参与其中,因为这种酶会抑制和稳定一些mRNA,或者使非编码RNA与mRNA结合,并阻止核酶结合。mRNA将被外核糖核酸酶或核糖核酸内酶所接触,从而迅速降解信使RNA。但是像Hfq或ProQ这样的蛋白质是非编码RNA调控所必需的。该蛋白具有多种功能,其最重要的功能是促进两个RNA分子之间的配对。
事实上,在金黄色葡萄球菌中蛋白质是保守的,且具有不同的功能。Rho蛋白不是必需的,它是毒力的衰减剂。葡萄球菌中不存在核内酶E,而是有两个外核糖核酸酶J1和J2。因此葡萄球菌的降解机制是不同的。Hfq存在于葡萄球菌中但不是RNA调控所必需的,因为这种蛋白质缺少一个阻止蛋白质识别RNA分子的特征,而ProQ在葡萄球菌中根本不存在。那么,与我们的新功能相关的关键RNA结合蛋白是什么呢?我们还未曾仔细研究葡萄球菌内部发生了什么。
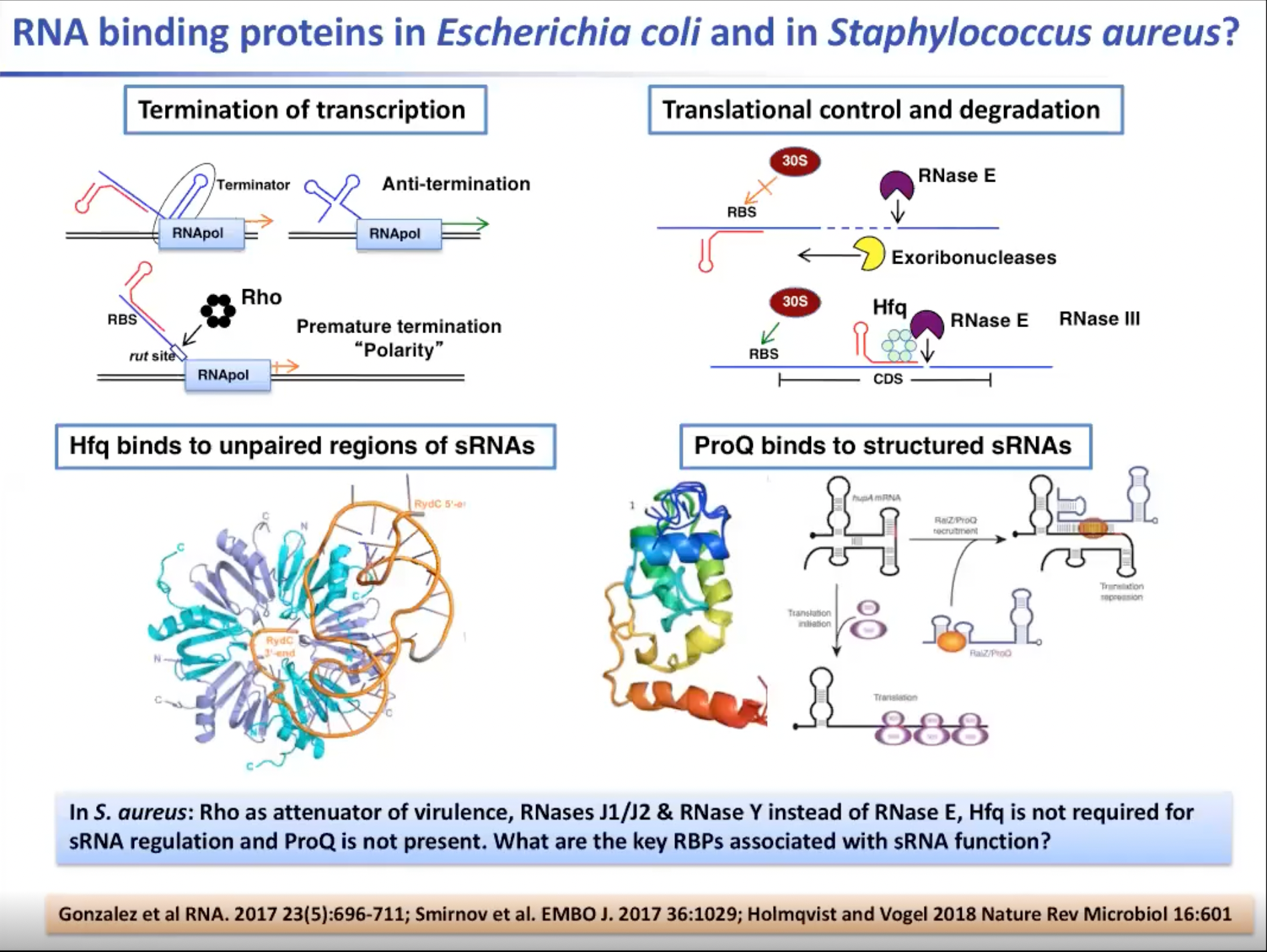
图3. RNA结合蛋白在大肠杆菌及金黄色葡萄球菌中有不同的功能
沿着亲缘树研究亲缘距离非常遥远的细菌是很重要的。因为基因调控机制不一定保守,小非编码RNA在远缘细菌中也不保守。它们只在密切相关的细菌中保守,如变形杆菌中的大肠杆菌和沙门氏菌在亲缘关系上非常接近,但大肠杆菌和金黄色葡萄球菌亲缘关系很远,它们有完全不同的小非编码RNA。
那么非编码RNA证实了物种特异性吗?这可能是事实,但它们调节共同的通路吗?在厚壁菌中,尤其是在葡萄球菌中,有哪些途径是由非编码RNA调控的?这都是我们希望解决的问题。
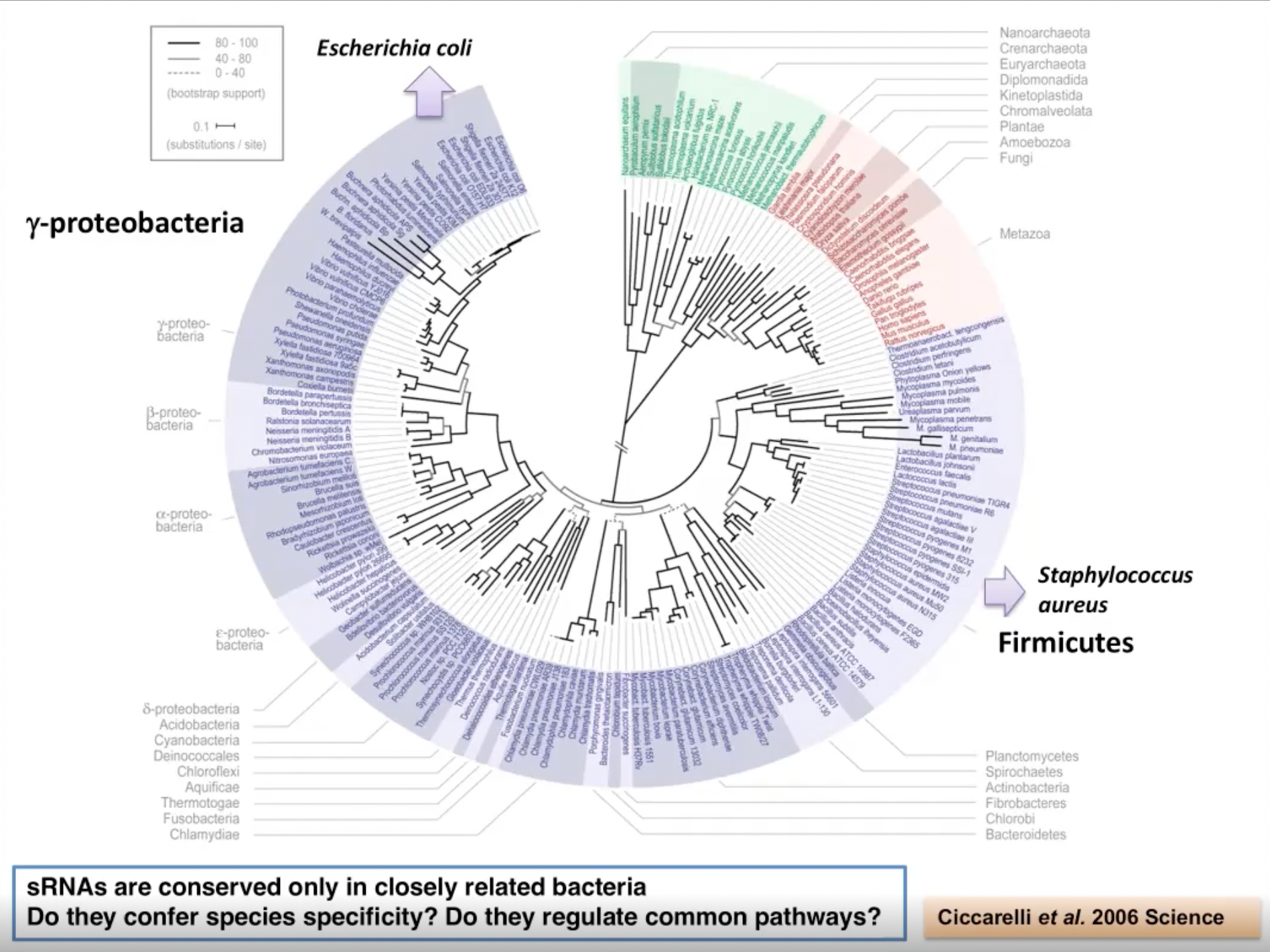
图4. 细菌的相对亲缘树研究
如我们所知,金黄色葡萄球菌是一种机会性产岩细菌。这是一种非常有趣的产岩细菌,因为葡萄球菌对人体器官不具有嗜性,但这种细菌能够感染人体基本上所有的组织和器官。它可能导致非常严重的感染,比如感染结核性肺炎或烟性肺炎。这是因为细菌产生了大量的变异。
葡萄球菌基因组中约10%是对变异因子的编码。这些变异因子涉及到添加机制和识别特定的人类蛋白质。一些变异因素阻止了细菌的变异机制,其他的会诱导生物信息,也会抑制生物信息。许多外毒素和外蛋白会针对免疫系统进行攻击。许多外毒素的活性这些毒素是细菌传播到宿主组织所必需的。
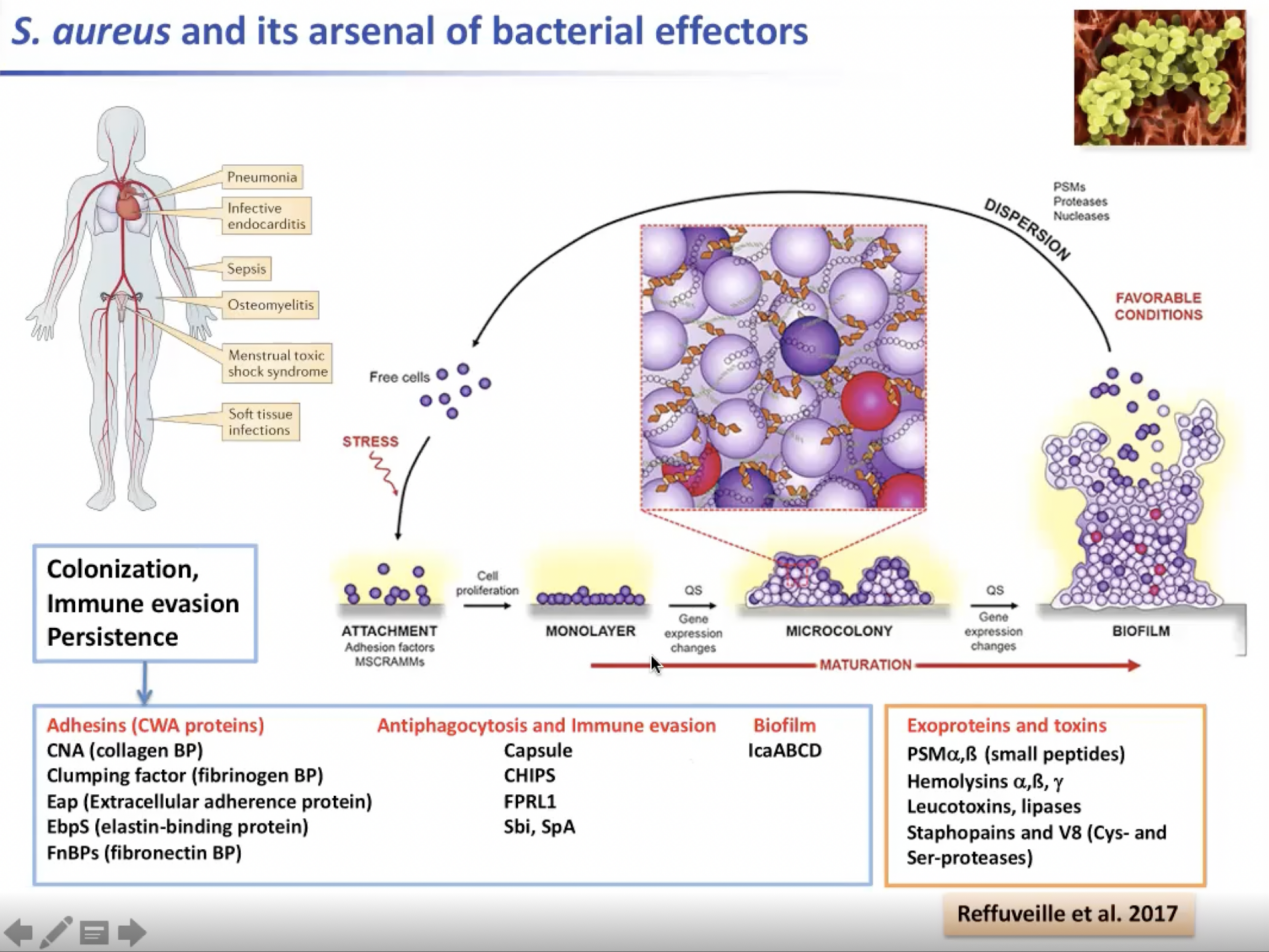
图5. 金黄色葡萄球菌及其效应因子
对葡萄球菌来说很有趣的一点是其黏附因子(Adhesion factors)受到严格的,暂时的调控。一个特定的毒力因子可以在不同层级上受到调控,可能是在转录层面,或在蛋白质层面受到调控。溶血素是一种可以被20种不同的调节因子调节的毒素。这些调节因子可能通过转录,RNA结合蛋白,调节RNA,或由双组分系统或是对代谢物作出反应的蛋白质得到。因此,多层调控使细菌能够对许多内部信号作出反应。
除此之外,临床分离株之间还存在差异。因为在临床分离的葡萄球菌菌株中,我们有不同的毒力岛(pathogenicity island),就会有不同的变异因子。其中一些还携带单核苷酸多态蛋白,因此只有一个突变可以消除方差因子的表达。因此,不同的临床分离株对人体的反应非常不同。这可能就是人体对由金黄色葡萄球菌引起的不同的传染性疾病有不同的反应的原因。
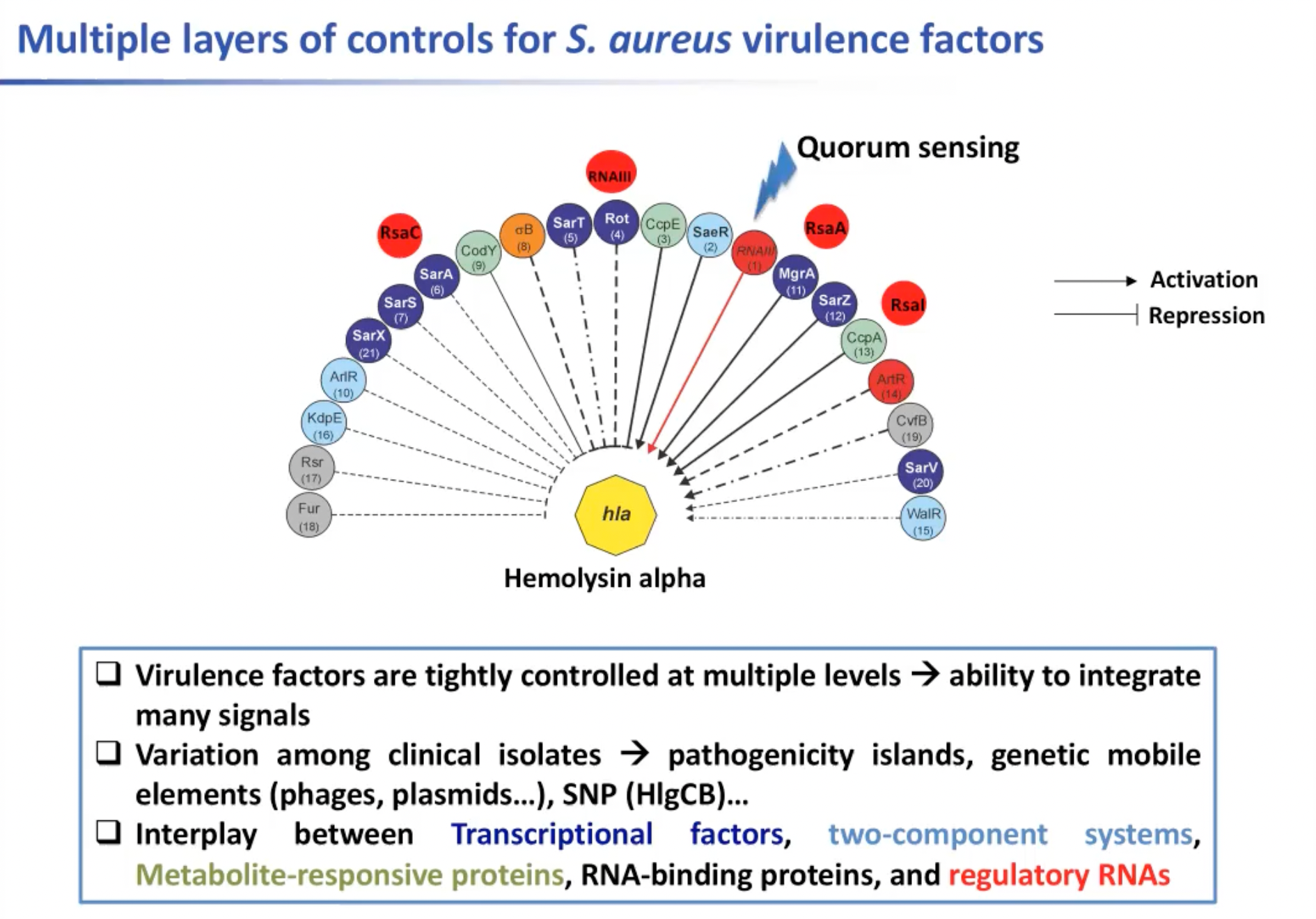
图6. 金黄色葡萄球菌的多层控制因子
但是在金黄色葡萄球菌中群体感应系统(Quorum Sensing System)对于毒力因子的表达是非常必要的。AGS系统编码小肽链,也就是argB这个小肽会从细菌外部分泌出来并由agrD蛋白的一个成员催熟,这个肽会随着细胞的生长而积累。当达到阈值浓度时,肽将与AgrC激酶膜结合,转移进入细胞。转移的这个激酶适合于反应调节因子AgrA。被激活的AgrA将自动激活它自己所在的肽链。许多参与新陈代谢的基因也参与调节。 一类外毒素,即一种较细的可溶性边缘肽,对人体细胞有系统活性。另外,AgrA会激活调节RNA的合成。这就是我们所说的RNAIII。
还有一种具有活性的被称为δ-溶酶基因的酚溶性调节素,图中被标注为红色的mRNA会直接充当附着RNA。该mRNA的3’端非常长,会特异性地与其他mRNA配对。现已知这种mRNA是通过编码毒力因子的方式参与免疫系统,并且是一种粘附因子。所以这两个变异因素对葡萄球菌来说是非常重要的。其中的机制很简单,这类RNA会隔离核糖体结合位点阻止核糖体与mRNA结合。mRNA会被双链核糖核酸酶III快速剪切,并且会迅速被破坏。但除此之外,调控RNAIII还会抑制另一个编码毒素抑制的mRNA。对毒素的抑制是由RNAIII的两个发夹基序与mRNA结合达成的。这个结合会被内切核糖核酸酶识别并切割,从而快速消灭编码毒素的mRNA。这个过程又会引入许多外毒素的合成。这个mRNA的5’ UTR还会与其他mRNA的UTR(或防止核酶结合的发夹结构)区域结合,以激活hla RNA的翻译过程。RNAIII与信使RNA的结合将阻止抑制结构,因此将允许核糖体与信使RNA结合并允许hla RNA的翻译。RNAIII同时会稳定翻译激活过程,这对调节mgrA RNA的翻译非常重要,mgrA RNA主导一种荚膜形成的活动,使细菌能够离开宿主,从而形成防御机制。
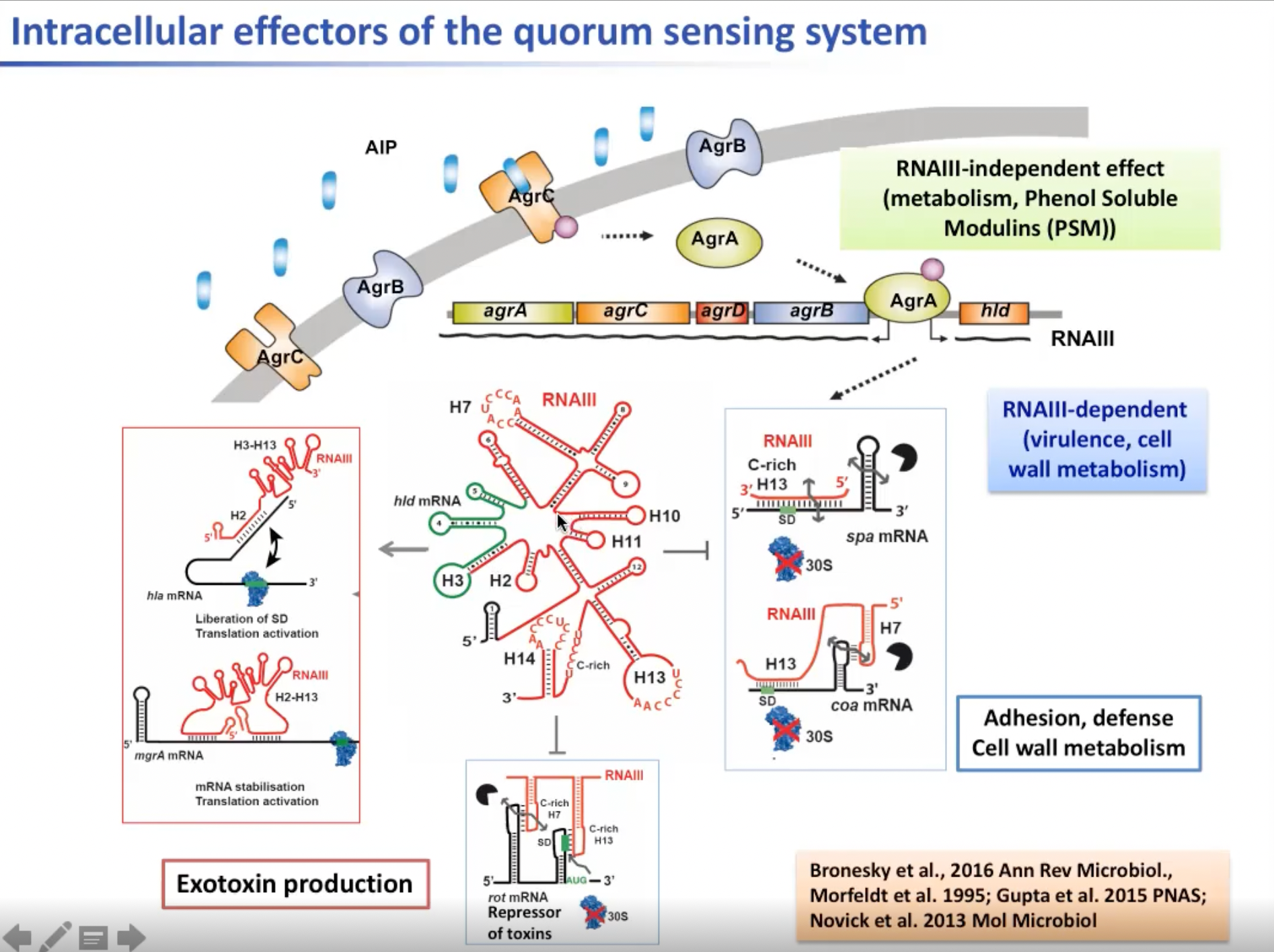
图7. 金黄色葡萄球菌胞内群体感应系统
总的来说,在整个过程中,RNAIII处于一个双前馈回路当中,通过调节转录会间接激活许多外毒素的合成,但同时也直接与mRNA结合生成降解毒素的hla mRNA。另外,RNAIII会抑制spa与coa的生成。因此,RNAIII既可以作为直接标记的翻译,也可以通过转录调节剂的调控间接作为转录和标记。有一个选择开关来调节基因表达。这是一个RNAIII的调节网络阐述了它是如何调节两组具有不同功能的基因的。然而,只有在信号持续的情况下,这种调节才能发生。这就是为什么RNAIII会随着它们的生长而强烈积累,并且一旦细菌生长,信号仍然是一致的。
但依然有很多问题尚未得到解答。比如RNAIII是否与蛋白质共同作用;RNAIII不仅对细胞的通常感应作出反应,同时也对许多环境信号作出反应,那么其调节网络是否与其他基因的网络相互连接;许多非编码RNA是如何参与到RNAIII的调节过程中的。
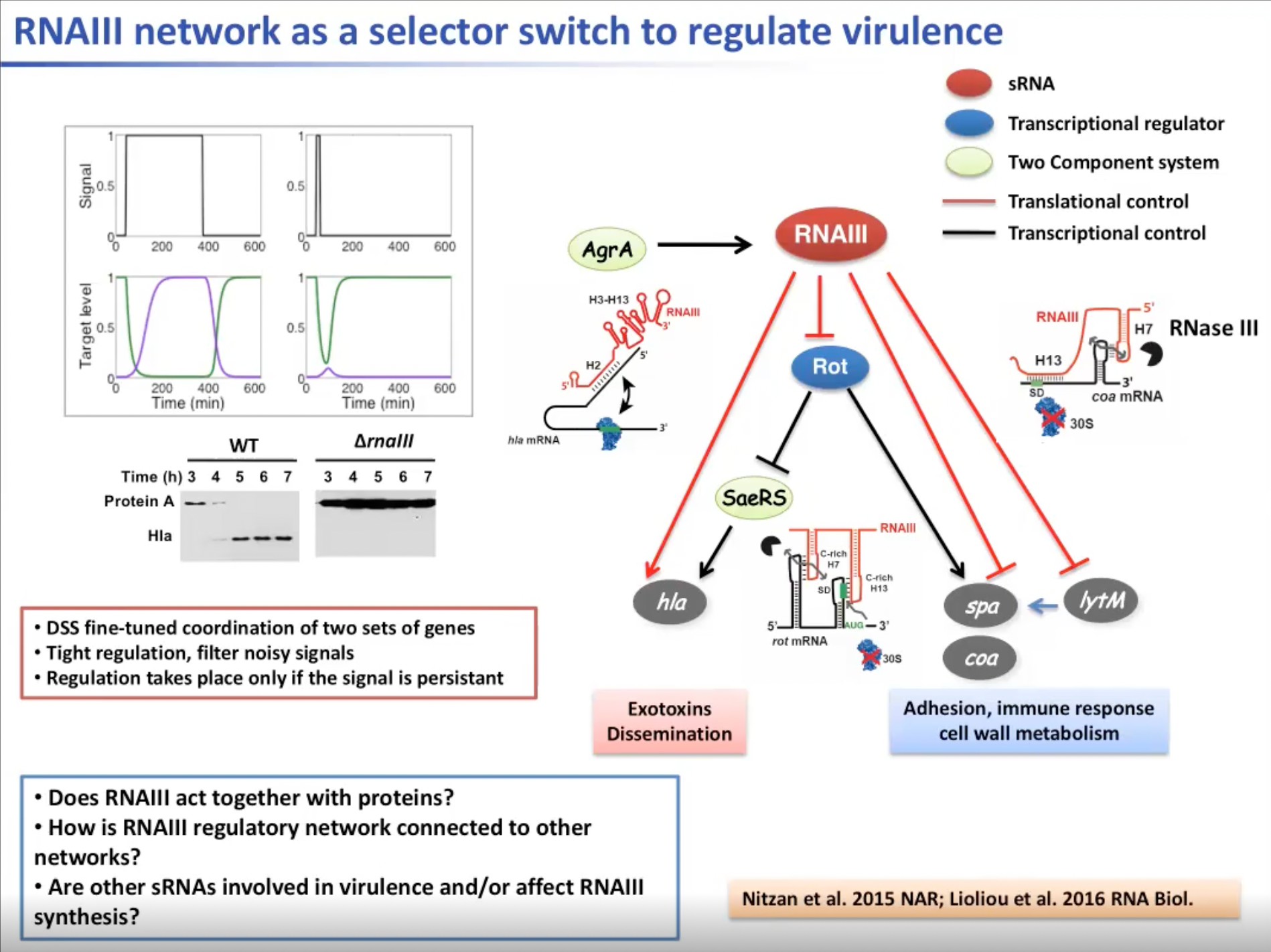
图8. RNAIII的调节网络
我们的第一个研究是与蛋白质复合的双链RNA,这是一种酶,是双链RNA的特例。我们可以证明核糖核酸内切酶III有很多不同的功能。它参与了RNA的转换。这种酶会破坏所有mRNA的结构,并参与到核糖体RNA的成熟,小非编码RNA的产生和mRNA稳定与加工等过程中。但其主要功能是通过识别许多不同的结构元素尤其是酶对mRNA进行反义调控,即被酶特异性识别以切割mRNA。这是一个可以阐述RNA结构对基因调控非常重要的例子。
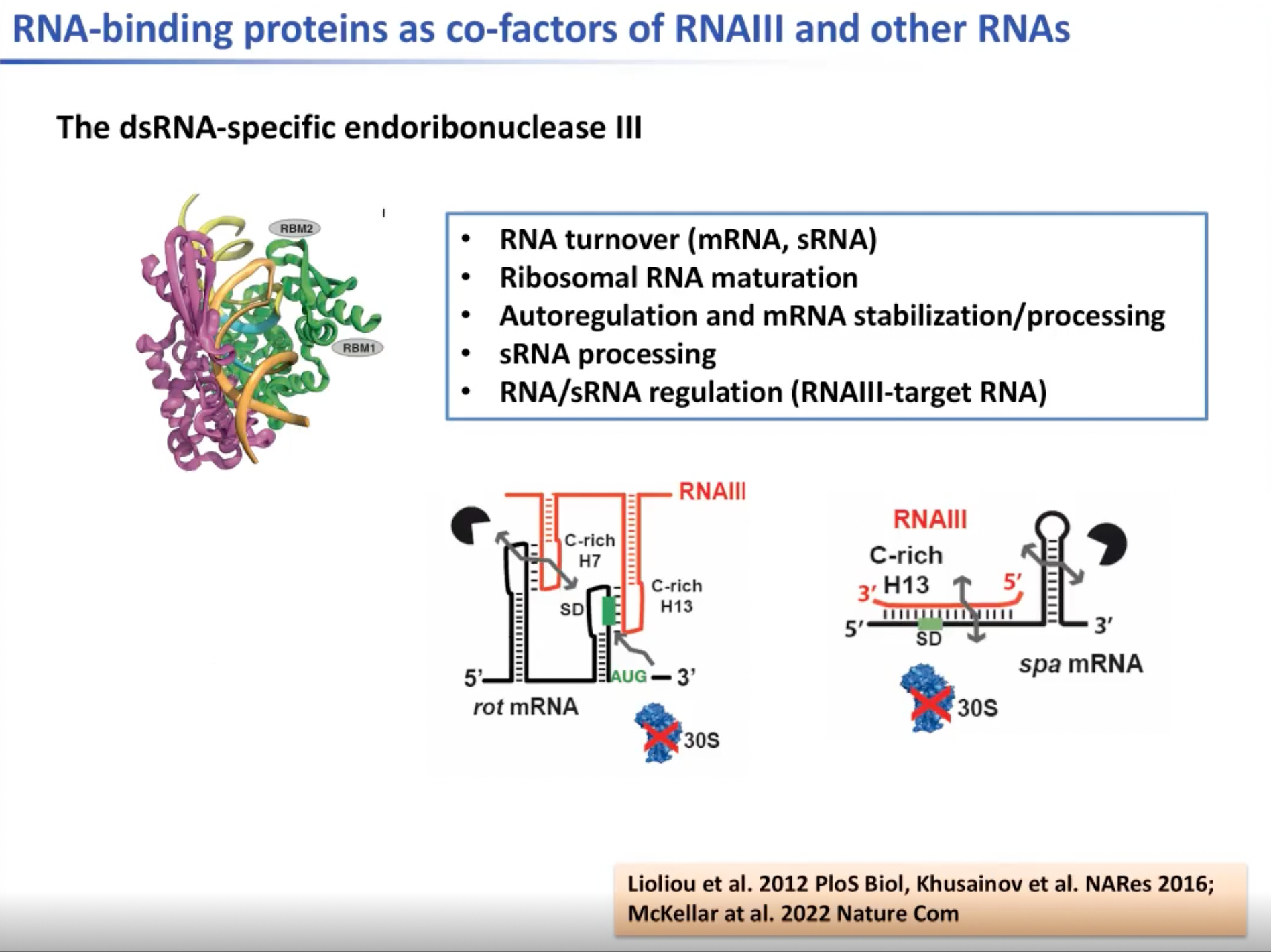
图9. 特例双链dsRNA--核糖核酸内切酶III
第二项研究是关于核糖体蛋白的。在大肠杆菌中核糖体蛋白S1与核糖体连接,这是大肠杆菌中核糖体起始过程所必需的。但在葡萄球菌中该过程不需要S1参与。该SauS1蛋白质缺少大肠杆菌中S1具有的两个与核糖体作用的区域。因此我们对S1的功能展开研究。我们对S1进行基于RIP-Seq的免疫沉淀反应测试。我们发现一组为PSM编码的mRNA和特定的sRNA会与之结合。
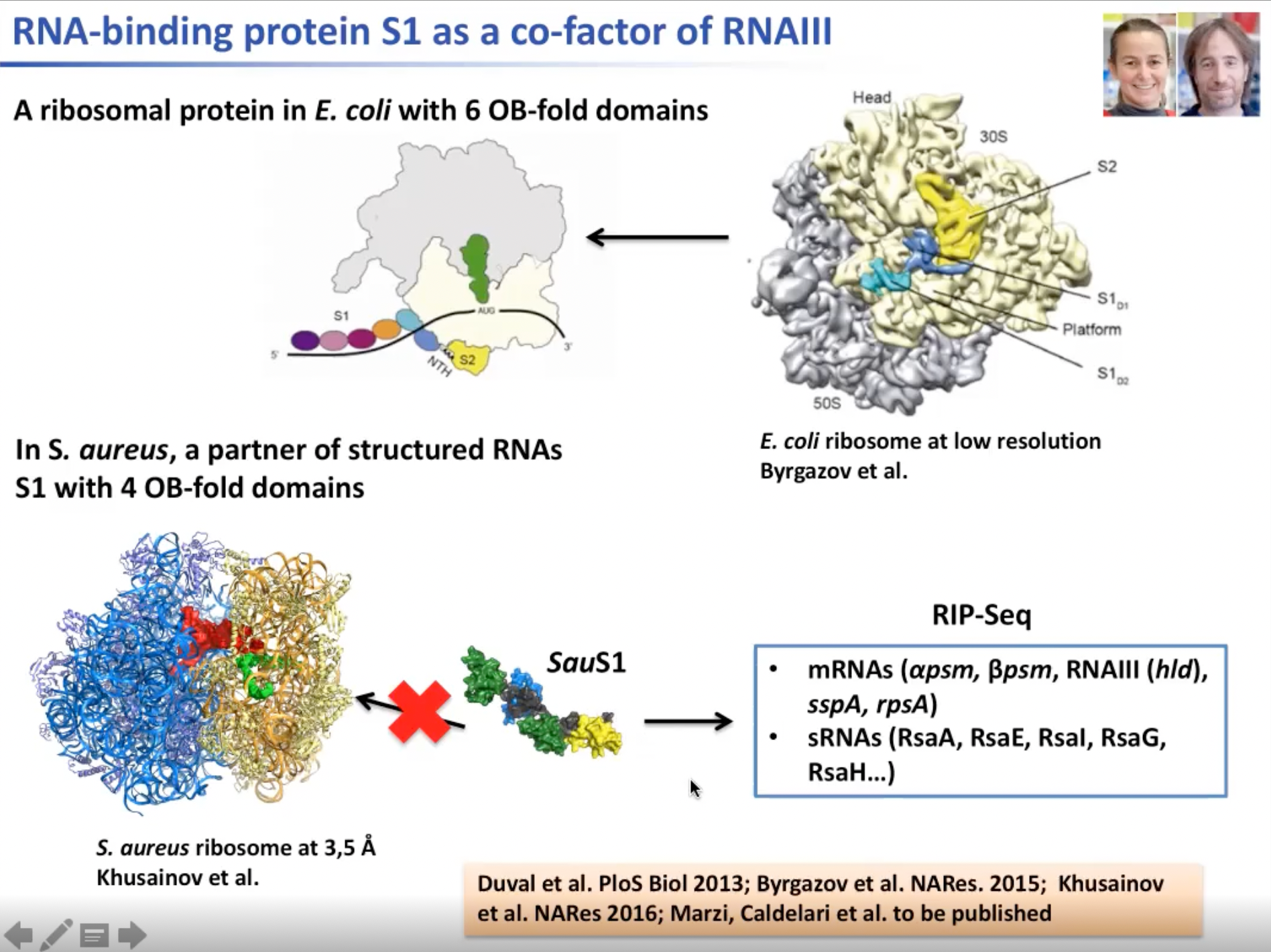
图10. 大肠杆菌中核糖体蛋白S1是核糖核酸内切酶III的co-factor之一
当不存在毒素特异性受体时,S1蛋白作用于细胞膜上,使一小块细胞膜解体并充当短期孔洞,这可使得很多酚溶性调节素(Phenol Soluble Modulins)进入细胞。因此S1对调节红细胞的细胞毒性与免疫系统非常重要。在实验中改变S1的量对小鼠进行腹膜内注射验证了这一点。因此S1是在葡萄球菌中发现的第一种可以改变毒力因子的RNA结合蛋白。
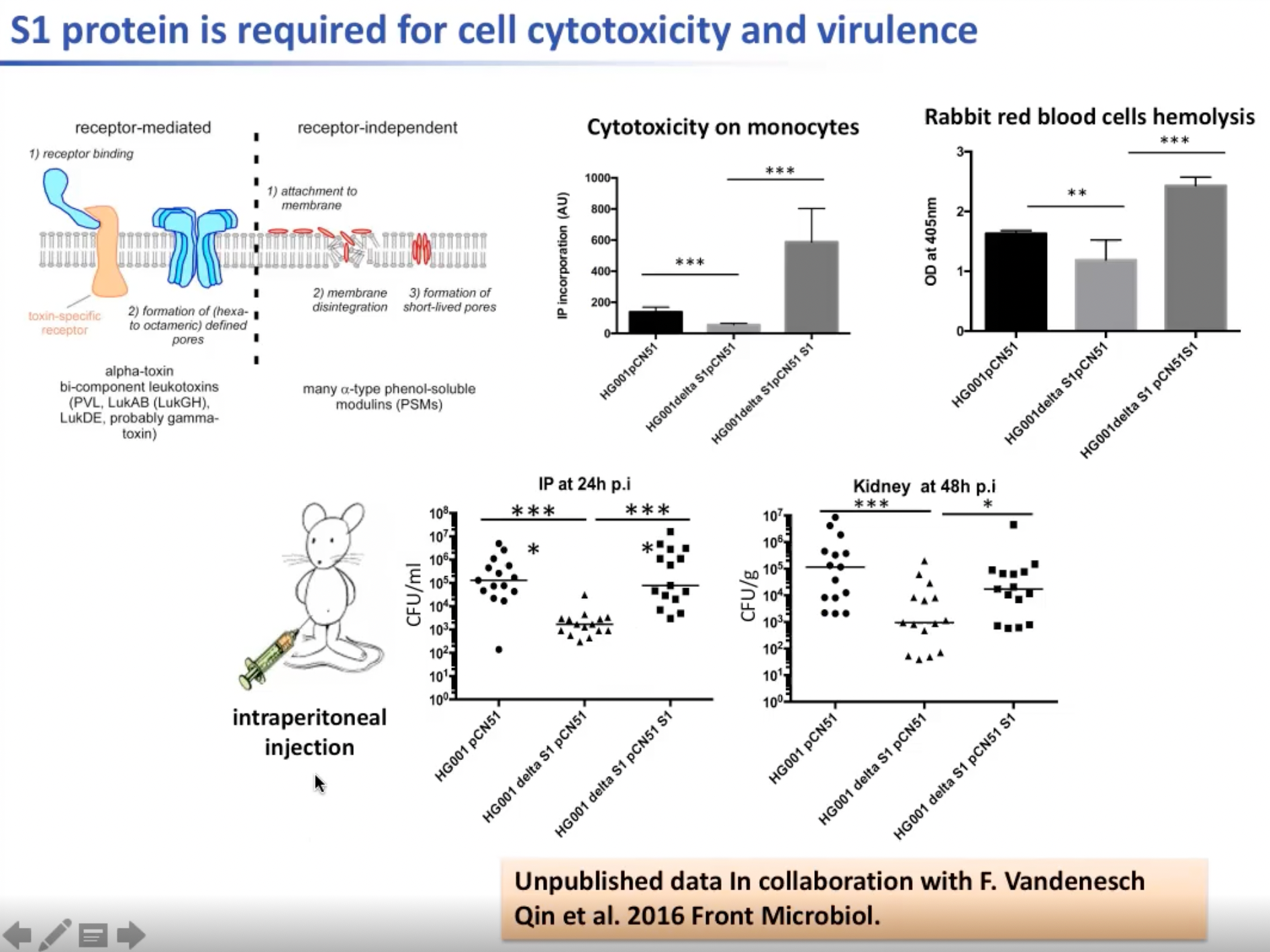
图11. S1蛋白对调节红细胞的细胞毒性与免疫系统非常重要
此处展示几个使用化学探测方法得到的编码PSM的mRNA。可以看到核糖体结合位点位于发夹弯结构之中。因此相比于RNAIII,发夹弯结构不同的mRNA没有被激活,从而无法产生PSM。所以上述的S1对mRNA的稳定是必要的。在S1减少的情况下,RNAIII与其他编码PSM的RNA的数量都会减少。
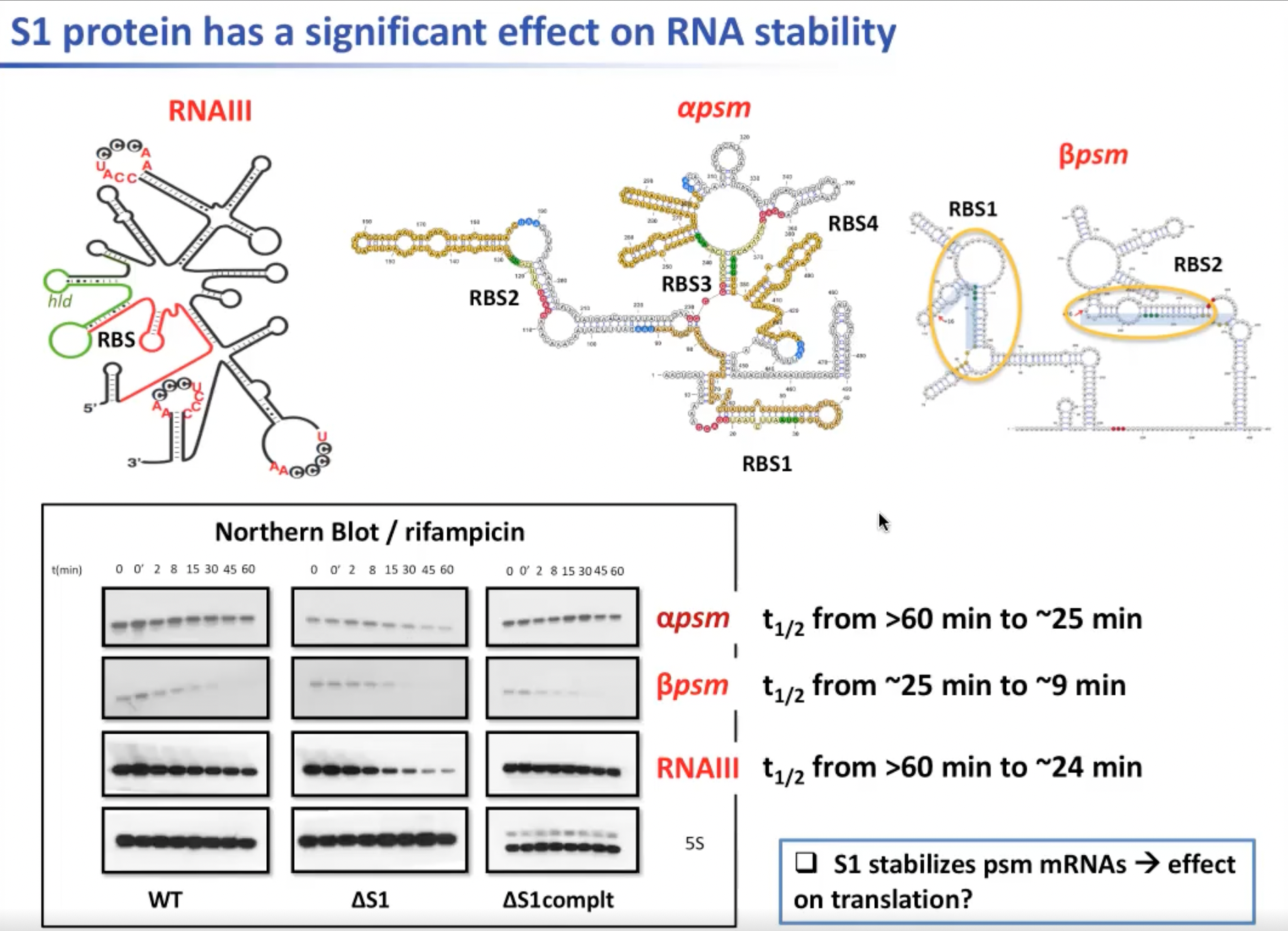
图12. S1蛋白对RNA稳定性的重要影响
但这个减少的过程不是直接的。我们追踪PSM在这类mRNA中的表达,发现截取其中一部分的表达,得到的PSM就不完整。实验中使用RNAIII的SauS1,tRNAi以及Sau30s RNA结合物与mRNA进行结合,会得到偏移16位的作为核糖体3’端的mRNA。如果缺失了上述的一部分mRNA,核糖体就无法高效地与RNAIII结合。因此与mRNA的结合会在构象上改变这类mRNA,使得核糖体可以与之结合。
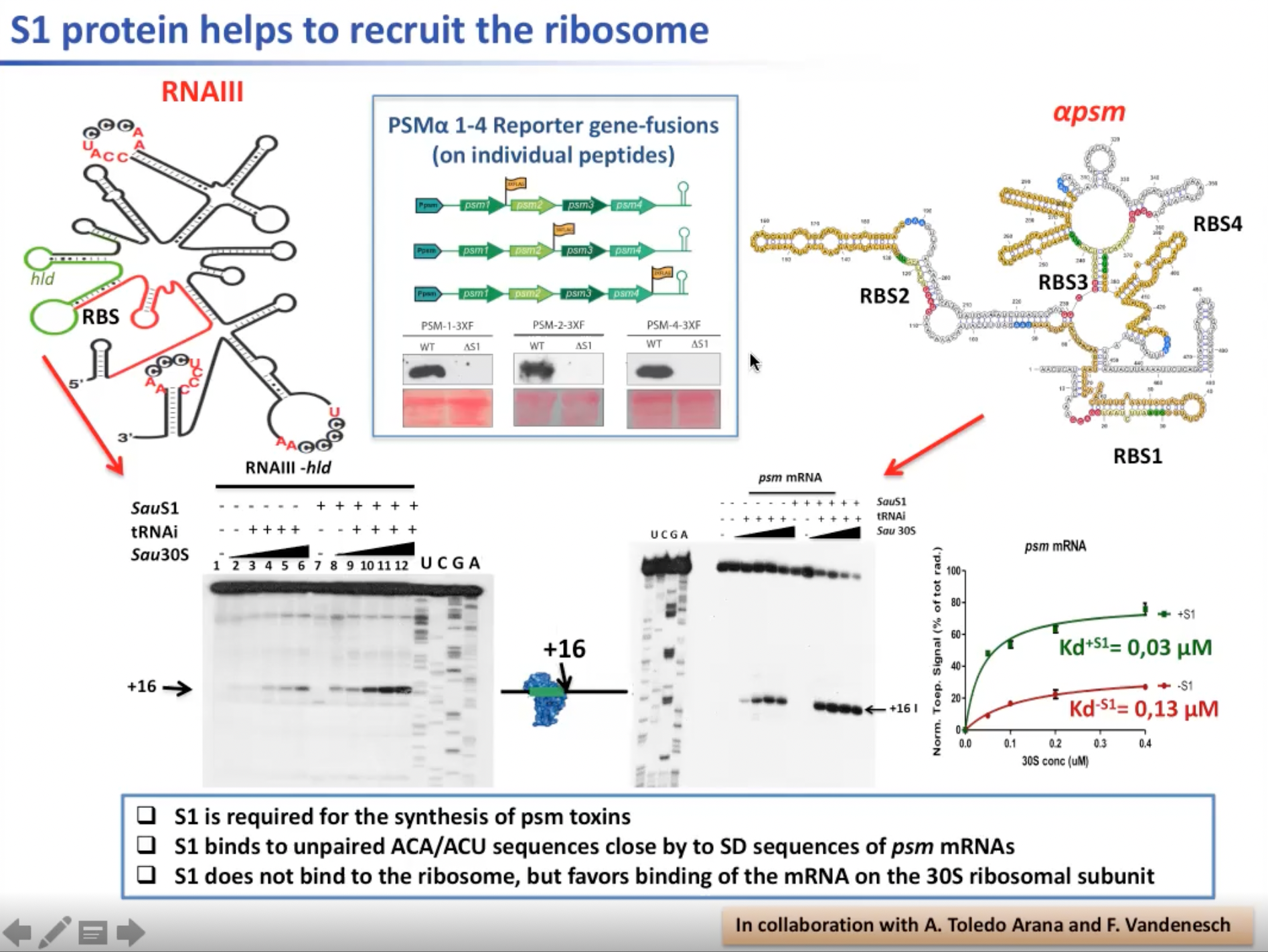
图13. S1蛋白帮助招募核糖体
实际上,一种名为陪护蛋白的蛋白质的四核结合位点可与mRNA结合,使得mRNA的构象稳定并展开便于与核糖体结合并翻译。因此需要S1与RNAIII一起调节一类毒素的合成。陪护蛋白对病毒因子的时间调节也很重要。同时RNAIII的转录也随着毒性的增长而增加。但其中RNAIII与Hld的量间有一段延迟,这段延迟可能由作为辅因子的S1产生。RNAIII产生的过程中,早期表达的毒性因子将被迅速抑制。但,RNAIII会随着浓度升高被用于生成外毒素。。
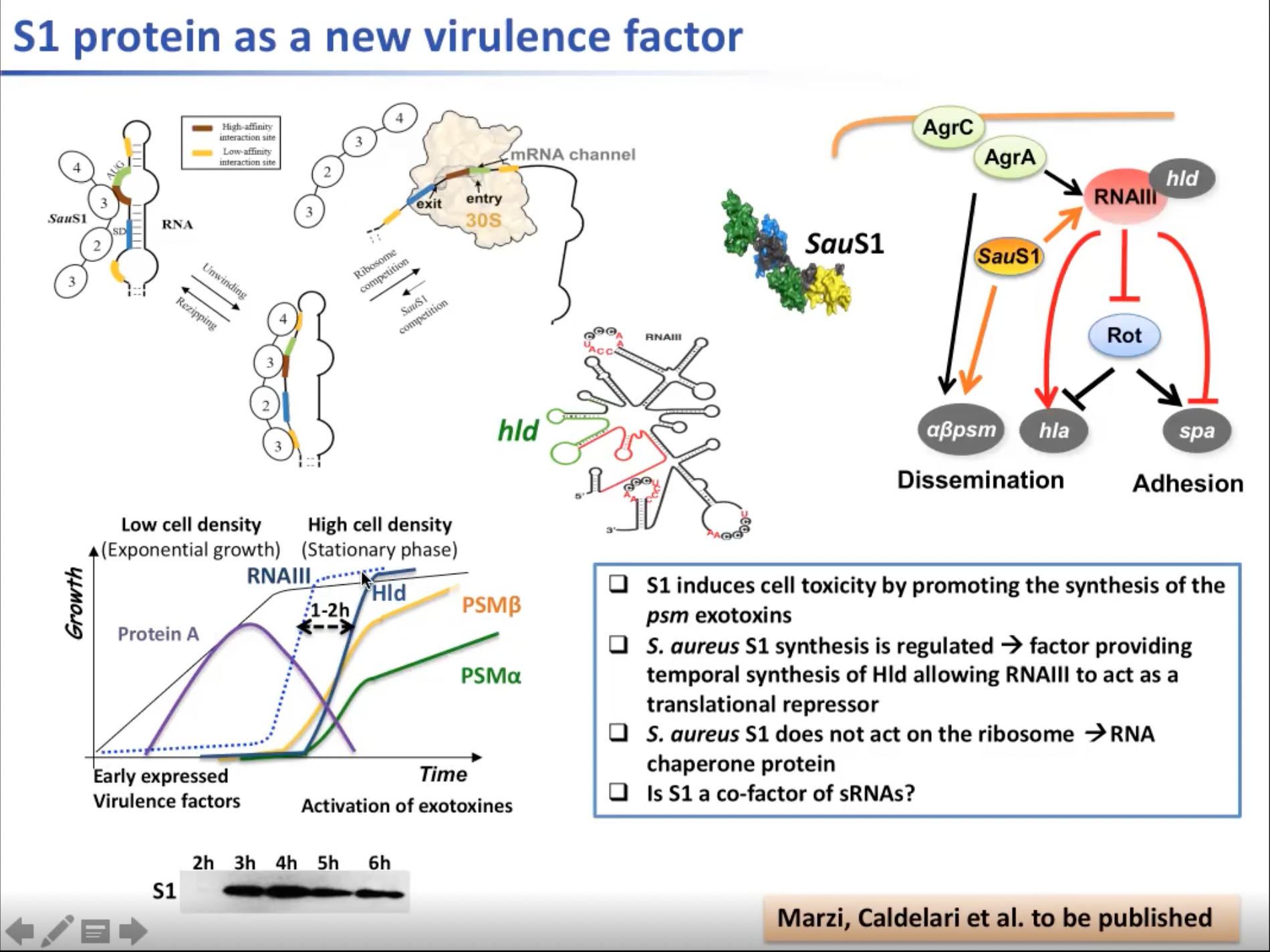
图14. S1蛋白是一个新的因子
因此RNAIII参与到一个复杂的调控网络之中。它直接作用于转录因子上来调节多种环境因子的合成,例如构建例如参与包膜形成的转录或调节因子MgrA等RNA时作为终止的Rho和Rot RNA,这些都是急性感染和细菌在宿主组织内传播所必需的。单个RNA分子与S1蛋白和RNase III一起能够调节30多个不同的基因。但正如一开始所说,RNAIII也能感知许多环境信号。事实上,很多转录都是调控因子它们发送不同的应激信号或碳源,比如GlcU,会直接与AGL系统相互作用并取代AGL系统。RNAIII的表达可能会激励,因此环境因子的表达要么会减少,要么会增加,因此可以很容易地通过转录或调节器来激发调节网络。
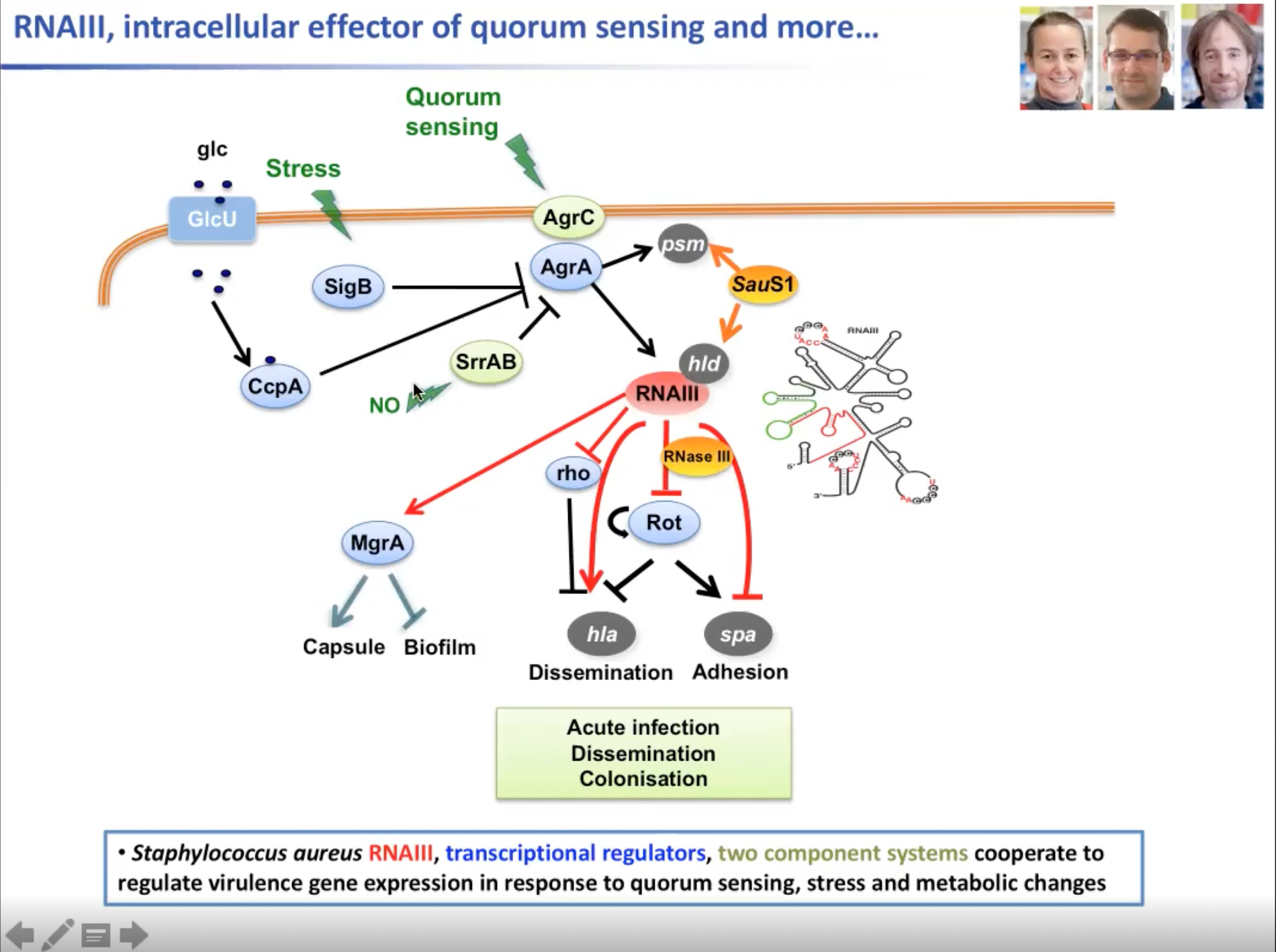
图15. RNAIII参与胞内复杂的调控网络
因此,人们认为,S1蛋白在大肠杆菌中是一种核糖体蛋白,而在葡萄球菌中是一种增强结构和短mRNA翻译的RNA结合蛋白,它同时也可能与小的非编码RNA调控有关。
小非编码RNA和大的RNA分子参与了代谢适应、毒力基因表达和金属稳态的十字路口,所有这些事件都是宿主保护相互作用所必需的。
另外,mRNA可以是小RNA的储存库,并且也有很多信使RNA编码转运蛋白的例子。研究还表明,一些非编码RNA携带有编码多肽的开放配对框,这就是RNAIII的定义。RNA修饰也与应激反应和毒力有关。

图16. 报告总结
接下来的工作:
MAPS和核糖体分析,以绘制特定sRNA的RNA靶组及其对翻译的影响;
研究RNA-RNA/RNP互作组在全局范围的相互作用(RIP-seq, CLASH, LIGR-seq, Grad-seq, SEC-Sec…),以对RNA互作组的深度学习进行研究;
在表转录组学中研究碱基修饰在解码中的作用与sRNA依赖性调节的联系,以应对应激和感染;
对RNA网络的动力学和生物学结果进行总结;
使用Dual-seg测量金黄色葡萄球菌和宿主细胞在感染情况下的全基因组转录变化;
在感染期间追踪RNA网络(动物模型);
研究微生物代谢物的影响。

图17. 未来研究方向
欢迎关注Guangzhou RNA club公众号、网站(rnaclub.rnacentre.org)、twitter(@RNA_club)。
 Guangzhou RNA club
Guangzhou RNA club